Xiaokang Wang1,
Kaihe Ye1,
Yanqing Lv1,
Songcheng Wei1,
Xiucun Li1,
Jinjin Ma1,
Xiaoqi Zhang2,
Chunling Ye1 ![]()
For correspondence:- Chunling Ye Email: yechunling2005@163.com Tel:+862085223843
Received: 14 September 2015 Accepted: 11 January 2016 Published: 28 February 2016
Citation: Wang X, Ye K, Lv Y, Wei S, Li X, Ma J, et al. Ameliorative effect and underlying mechanisms of total triterpenoids from Psidium guajava Linn (Myrtaceae) leaf on high-fat streptozotocin-induced diabetic peripheral neuropathy in rats. Trop J Pharm Res 2016; 15(2):327-333 doi: 10.4314/tjpr.v15i2.15
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate whether the total triterpenoids extracted from Psidium Guajava leaves (TTPGL) attenuate the development of diabetic peripheral neuropathy in rats by regulating the NF-κB pathway of the inflammatory process and its signaling mediators.
Methods: All the Sprague Dawley rats used were maintained in a clean environment on a 12 h light/12 h dark cycle. High-fat feeding and intraperitoneal injection of 40 mg/kg streptozotocin (STZ) were used to induce diabetes in the rats. The rats were randomly divided into 5 groups: diabetic mellitus (DM) group; TTPGL - 30 group, TTPGL - 60 group and TTPGL - 120 group treated by intragastric administration (i.g) with 30, 100 and 120 mg/kg/day TTPGL, respectively. The well-established drug, rosiglitazone (RSG, 3 mg/k/d, i.g.), was used as positive control. Normal rats served as control group. Nerve conduction velocity and sensitive tests were measured on weeks 1, 4 and 8. After 8 weeks administration, ex
Results: Compared to DM group, TTPGL (especially 120 mg / kg dose) treatment improved (p < 0.05) physical functions and provided neuronal protection in high - fat/streptozotocin - induced peripheral neuropathy rats. We found that the ex
Conclusion: TTPGL offers a potential therapeutic approach for the treatment of diabetic peripheral neuropathy.
Introduction
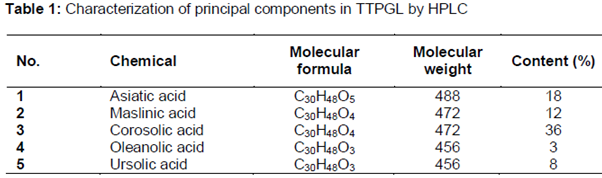
Psidium guajava Linn., commonly known as guava, has been naturalized in South - East Asia [1]. In oriental regions, such as Taiwan, guava leaves is boiled and the extract is used in folk medicine for the treatment of diabetes [2] but few of the extract has been investigated for their preclinical or clinical efficacy. Psidium guajava Linn. (Myrtaceae) is a small evergreen tree and its leaves are short-petiolate with the blade oval with prominent pinnate veins. Leaves of this plant have been reported to contain several compounds such as triterpenoids [1] and flavonoids [3]. Recent tropical relevance studies have shown that the leaves of Psidium guajava have been used for the treatment of a number of diseases, e.g. as an anti - inflammatory, for diabetes, pain relief and anti - pyretic. Moreover, previous phytochemical investigations had demonstrated that triterpenoids are the principle compounds isolated from the leaves [4]. As a continuing research of this potential drug for diabetic complications, we conduct this study on 5 known triterpenes: Asiatic acid, Maslinic acid, Corosolic acid, Oleanolic acid, and Ursolic acid.
Many of the pathophysiological pathways of diabetic neuropathy focused on inflammation and its mechanisms appear to converge upon the activation of NF - κB in diabetic neuropathy [5,6]. In addition to modulating of pro - inflammatory responses, NF - κB causes the transcription of pro - inflammatory mediators such as interleukin - 6 (IL - 6), tumor necrosis factor - α (TNF - α) and inducible nitric oxide synthase (iNOS) [7]. It is reported that Asiatic acid and Oleanolic acid possess anti - inflammatory activities [8,9]. Therefore, it is essential to investigate whether TTPGL has protective effect on DPN by suppressing NF - κB pathway and thus inhibits pro - inflammatory mediators.
Methods
Chemicals and reagents
Protein kinase B (Akt), inhibitory κB kinase alpha (IKKα) and phosphor - IKKα and NF - κB - p65, horse radish per - oxidase (HRP) - conjugated antirabbit IgG were purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA). Other reagents were purchased from Sigma - Aldrich (St. Louis, MO, USA) unless indicated elsewhere.
Extraction and isolation compounds of TTPGL
The leaves of Psidium guajava were collected from their natural habitat in Guangzhou, Guangdong, in October of 2007. The plant was authenticated by Prof. Shu - Yuan Li (Guangdong Pharmaceutical University, Guangzhou, China). A voucher specimen (no. 2007101801) was deposited at the herbarium of Institute of Traditional Chinese Medicine & Natural Products, Jinan University.
The air-dried leaves of P. guajava (5.0 kg) were powdered and percolated with 85 % (v/v) EtOH at room temperature for three times (3 × 35 liters). After removing the solvent, the alcohol extract (110 g) was suspended in H2O and partitioned successively by petroleum acetone (PA) to obtain the PA extract (88 g) and water extract (22 g), respectively. The PA extract (88 g) was separated by silica gel (200 - 300 mesh) CC and eluted with TFA - MeOH to obtain fractions. Fractions were separated by silica gel column with gradient mixtures of MeOH - TFA as eluents to yield subfractions. The PA extract (88.0 g) was separated by silica gel (200 - 300 mesh) CC and eluted with MeOH - TFA to obtain 8 fractions (C1 - C8). Fractions were subjected to silica gel column eluted with mixtures of MeOH - TFA (8500:1; v/v) to yield compounds 1 (120 mg, 98 %), 2 (120 mg, 98 %), 3 (120 mg, 98 %), 4 (150 mg, 98 %) and 5 (200 mg, 98 %).
Induction of hyperglycemia in rats and blood glucose measurement
Adult male Sprague Dawley rats (130 - 140 g, animal quality certificate number SCXK 2013 - 0002) were maintained under standard condition (a constant temperature on a clean environment in a 12 h light/12 h dark cycle). All procedures were carried out in accordance with the principles of U.S. National Institutes of Health guidelines [10]. All study protocols were approved by the local government committee for animal research (approval ref no. 20140110103118). The rats were fasted for 12 h prior to injection, rats were injected with STZ (40 mg/kg, dissolved in 0.1 mmol/L sodium citrate buffer, pH 4.5) intraperitoneally, while the control group rats were injected with the vehicle citrate buffer. One week later, the rats with blood glucose above 11.1 mmol/L were considered diabetic and selected for further pharmacological studies [11]; secondly, animals were fed their respective diets until the end of the study as described: diabetic rats were fed with a high - fat diet (consisting of 70 % standard laboratory diet, 15 % carbohydrate, 10 % lard and 5 % yolk powder) while the healthy rats were fed with normal chow. Blood samples obtained from the tail vein of rats were monitored for glucose once a week using reagent strips with a glucometer (Onetouch Ultra 2, Johnson & Johnson, Milpitas, CA, USA).
Experimental animal groups
Diabetic rats were randomized into 5 groups of 10 animals each and were still fed with a high - fat diet, namely DM (untreated diabetic rats) group, TTPGL - 30 (30 mg/kg/d, i.g.) group, TTPGL - 60 (60 mg/kg/d, i.g.) group, TTPGL - 120 (120 mg/kg/d, i.g.) group and RSG (3 mg/kg/d, i.g.) group (as a hypoglycemic positive control). Normal rats served as control group (n = 10). 8 weeks were designed for each group. The samples from the rats were collected during anesthesia and finally the rats were sacrificed.
Nerve function measurement
We measured MNCV and SNCV in all rats at 1, 4 and 8 weeks after treatment in rats. MNCV was calculated by subtracting the distal latency from the proximal latency, and recorded in meters per second as the ratio of the distance in millimeters. SNCV was measured in the digital nerve to the second toe, using the smallest intensity current that resulted in an amplitude response, and was recorded in meters per second as the ratio of distance between stimulating and recording electrodes [12,13].
To determine the thermal response latency, rats were placed within a plexi glass chamber and allowed to acclimatize for at least 20 min. A thermal stimulation meter from ChengDu Technology & Market Co., LTD. (Chengdu, China) was used. The device was activated after placing the stimulator directly beneath the tail. Tail flick latency in response to the radiant heat (a heating rate of 1.3 °C per second, cut - off time of 16 s for rats) was recorded. At least three readings per rat were taken at 15 min intervals, and the average was calculated.
Immunofluorescence (IF) staining
We fixed and embedded sciatic nerves in paraffin with transverse section. The paraffin sections, cut at 5 μm, were deparaffinized in xylene and rehydrated in a series of graded alcohols to distilled water. Antigen retrieval was then performed in 10 mmol/L citrate, pH 6.0, using 1100 - W microwave oven for 3 min at high power and 12 min at 50 % power. The slides were allowed to cool at room temperature. Tissues were then incubated for 10 min in 0.3 % hydrogen peroxide to block endogenous peroxidase activity. The slices were then washed for 5 min in PBS and were incubated in 0.1 % Triton X - 100 and 5 % goat serum for 30 min. Tissues were then incubated with primary monoclonal antibody raised against IL - 6 (1:100), TNF (1:100), iNOS (1:50) overnight at 4 °C in a humidified chamber. Following extensive washing, the slices were incubated with fluorescein conjugated secondary antibody at room temperature for 1.5 h [14]. The nuclei were counterstained using 4’ - 6’diamidino - 2 - phenylindole (DAPI). All immunofluorescence staining were performed in replicates on 3 sciatic nerves. The sections were examined under a Zeiss Axiophot fluorescence microscope. To examine inflammatory cytokines infiltration in the sciatic nerve, randomly 15 - 20 fields or focuses from each animal were observed and analyzed by Image J (version 1.46 m) software.
Western blotting assay
Protein lysates were obtained by homogenizing sciatic nerves with lysis buffer containing 1 % Triton X - 100, 150 mmol/L NaCl, 1 mmol/L EDTA, 2.5 mmol/L sodium pyrophosphate, 1 mmol/L β - glycerophosphate, 1 % Na3VO4, 1 μg/mL leupeptin, 1 μg/mL aprotinin and 20 mmol/L Tris (pH 7.5). Equal amounts of 40 μg proteins were separated by SDS - PAGE (10 %) and transferred to a nitrocellulose membrane (Pall Life Sciences, Florida, USA). After blocking with 3 % bovine serum albumin, membranes were incubated with primary antibodies against Akt, IKKα, NF - κB - p65, phosphor - Akt, phosphor - IKKα, and phosphor - NF - κB - p65 for 12 h at 4 °C. After washing, membranes were then incubated with anti - rabbit IgG conjugated to HRP (1:1000 dilution) at room temperature for 1 h. Membranes were washed thrice, visualized with Immobil on Western HRP Substrate, and scanned with a Kodak Digital Imaging System (Gel Logic 2200 Pro; Kodak, Tokyo, Japan). β - actin expression was set as internal control and the relative band densities were quantified by densitometry.
Statistical analysis
Statistical analysis was performed using SPSS 19.0 software (SPSS, Chicago, USA). Data are expressed as mean ± SEM. Multiple groups were compared using one - way analysis of variance followed by Tukey HSD post hoc comparison. P < 0.05 was considered statistically significant.
Results
The structures of compound 1 - 5 were identified by comparing the 1H NMR and 13C NMR spectral data with previous literature reports. Principal components are shown in .
Effect of TTPGL on fasting blood glucose (FBG)
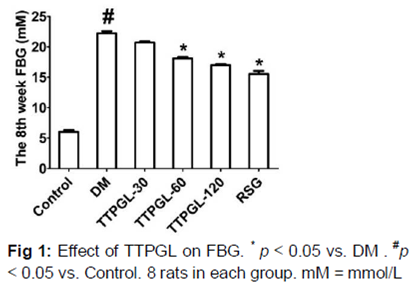
Levels of FBG in serum are shown in . It showed significant differences (p < 0.05) in the TTPGL - 60, TTPGL - 120 and RSG groups compared to the DM group.
Effect of TTPGL on neurological function
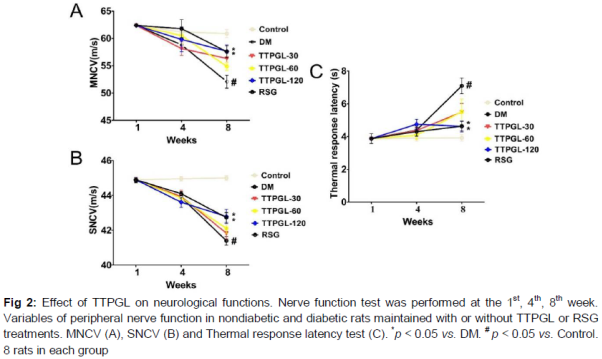
The definition of DPN may include a focused neurological examination or nerve conduction studies, or, all of these [5]. As shown in , within 8 weeks of the onset of diabetes induced by STZ, a severe peripheral neuropathy developed. Rats with 8 - week duration of high - fat/STZ diabetes had development of MNCV (16 %) and SNCV (9 %) deficits compared with Control. In contrast, after 8 weeks of TTPGL - 120/RSG treatment, all nerve conduction velocities demonstrated a marked improvement (p < 0.05) (A and B). In addition, compared to DM group, treatment of TTPGL - 120 and RSG groups markedly (p < 0.05) produced a reduction in thermal latency (C).
Effect of TTPGL on sciatic nerve TNF - α, IL - 6 and iNOS expression
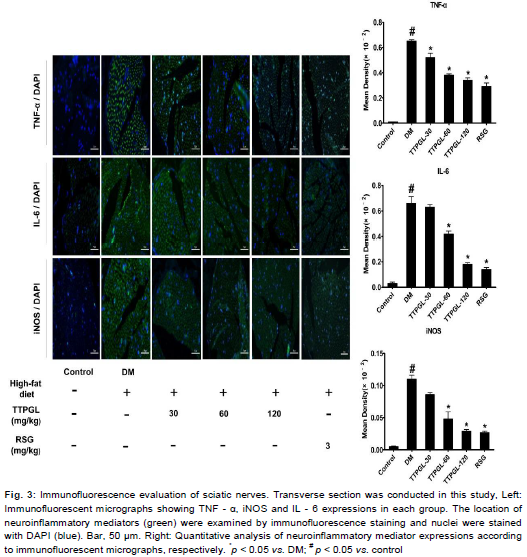
Based on findings above, using IF staining, we confirmed the inhibitory effect of TTPGL on inflammatory mediators. Compared to the Control, DM group exhibited significant (p < 0.05) increase in fluorescence intensity, while treatment with TTPGL decreased the fluorescence intensity. Those micrographs indicated that TTPGL inhibited the production of the inflammatory mediators TNF - α, IL - 6 and iNOS at the dose of 60 or 120 mg/kg, respectively.
Quantitative analysis of inflammatory expressions are shown in . RSG effects on high - fat diet/STZ induced diabetic rats is similar to that of TTPGL (60 or 120 mg/kg). These data are in accordance to our previous hypothesis that TTPGL could be developed into a protective agent against NF - κB - mediated expression of neuroinflammatory mediators.
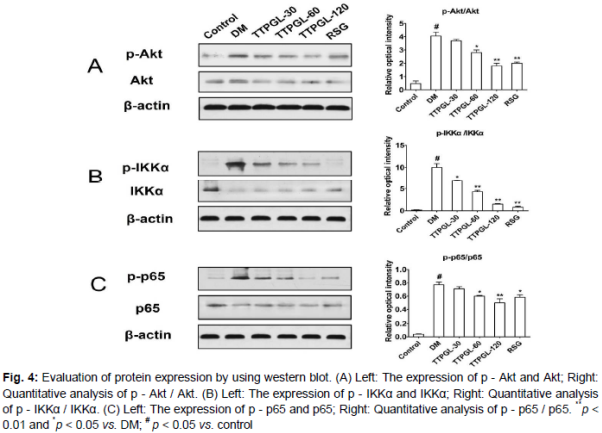
Effect of TTPGL on NF - κB activation via the Akt/IKKα pathway
As shown in , the experiments revealed that the constitutive NF - κB cascade in sciatic nerve of diabetic rats was significantly (p < 0.05) increased when compared with that of the Control. TTPGL dose - dependently decreased (p < 0.05 or p < 0.01) the phosphorylation levels of Akt (A), IKKα (B) and which subsequently suppressed phosphorylation levels of NF - κB - p65 (C). RSG as well as TTPGL (60 mg/kg, 120 mg/kg) significantly (p < 0.05) inhibited the phosphorylation levels of Akt, IKKα and NF - κB - p65 than that of the DM group (). Furthermore, TTPGL at dose of 120 mg/kg exhibited extremely significant (p < 0.01) decrease in protein expression. These findings suggested that TTPGL suppresses the NF - κB - p65 activation and regulate NF - κB - p65 activity through Akt/IKKα pathway in a dose-dependent manner.
Discussion
The findings reported herein show that long - term TTPGL treatment exerted protective effects in high - fat/STZ relevant model for DPN in vivo. Through a combination of biochemical and histological results, TTPGL plays a role in suppressing the expression of inflammatory mediators. Mechanistic studies suggested that TTPGL significantly ameliorates DPN through negatively regulating Akt/IKKα mediated NF - κB pathway.
In present studies, treatment with TTPGL - 120 ameliorated the deficits in MNCV, SNCV versus the DM group. The sensorimotor alteration, is one of the debilitating features of diabetic neuropathy in which nerve sensitization and inflammation are the commonly observed mechanisms [15,16]. The favorable effects of TTPGL on nerve - functional deficits may be due to its lightly hypoglycemic effect and its effect on inflammatory infiltration. The exact principle responsible for the hypoglycemia and amelioration effect on DPN were not investigated in the current study. Moreover, further studies would be required to determine the exact mechanism(s) and site of action of the extract.
Inflammatory cytokines which have been observed to contribute to the development of neuropathy and neuropathic pain in diabetes includes TNF - α and IL - 6 [17]. Treatment with TTPGL was able to attenuate the increased levels of TNF - α and IL - 6 in treated group which might have aided in decreasing inflammatory damage found in sciatic nerves. It is known that NF - κB activation induces the expression of iNOS, an enzyme induced during inflammatory conditions which specifically binds to COX-2 and S - nitrosylates to enhance COX - 2 activity [18]. TTPGL - 120 treatment inhibited the induction of iNOS which may have further decreased COX - 2 activity as well as inflammatory damage in neurons.
In this study, we observed TTPGL ameliorative effect and investigated correlated underlying mechanisms in the diabetes peripheral neuropathy rats. The NF - κB signaling cascade interacts with several parallel pathways including the signaling cascades initiated by phosphatidylinositol 3 - kinase PI3K and Akt [19,20]. Akt, functioning as a key downstream target of PI3K, has been suggested to function as an IKK (IκB kinase) kinase [21]. Western blot analysis showed that the expressions of p - Akt and p - IKKα in the DM group were significantly higher (p < 0.01) than those in the Control. In addition, the increased expressions of p - Akt and p - IKKα after STZ treatment could be attenuated by TTPGL administration. These results were indicated that TTPGL exerts ameliorative effect in DPN rats associated with changes in NF - κB cascade.
Conclusion
The findings provide the first direct evidence that TTPGL effectively decreased the release of inflammatory mediators in vivo. The efficacy of TTPGL, at least in part, may be due to the combined modulation of levels of NF - κB and inflammatory mediators as seen in our study. Interactions among TTPGL and other inflammatory mediators implicated in the pathogenesis of DPN also need to be explored. However, this work provides rationale for the evaluation of TTPGL in easily accessible natural sources as agents for diabetic complicates.
Declarations
Acknowledgement
References
Archives


News Updates